Chemically induced cuticle mutation affecting epidermal conductance to water vapor and disease susceptibility in Sorghum bicolor (L.) Moench
1
1994
... 植物一生固着于地面,因而更易遭受生物和非生物逆境胁迫.在长期进化过程中,植物从形态结构、生理生化等诸多方面演化形成一套完整的适应机制以抵御逆境胁迫.叶片作为地上部感知外界环境最敏感器官,其形态结构表现出与环境之间良好的适应协同性,如叶片表面蜡质可以抵御病原微生物和虫害以及其他生物胁迫和非生物胁迫[1~2],表皮腺体分泌物对生物胁迫和非生物胁迫[3],角质膜调节非气孔性蒸腾与体表温度[4]和防御病虫害叮咬[5~6]等.气孔是植物叶片特有的结构,在调节植物与环境之间气体交换、水分扩散中发挥重要作用. ...
The effect of the environment on the permeability and composition of citrus leaf cuticles:Ⅱ.composition of soluble cuticular lipids and correlation with transport properties
1
1990
... 植物一生固着于地面,因而更易遭受生物和非生物逆境胁迫.在长期进化过程中,植物从形态结构、生理生化等诸多方面演化形成一套完整的适应机制以抵御逆境胁迫.叶片作为地上部感知外界环境最敏感器官,其形态结构表现出与环境之间良好的适应协同性,如叶片表面蜡质可以抵御病原微生物和虫害以及其他生物胁迫和非生物胁迫[1~2],表皮腺体分泌物对生物胁迫和非生物胁迫[3],角质膜调节非气孔性蒸腾与体表温度[4]和防御病虫害叮咬[5~6]等.气孔是植物叶片特有的结构,在调节植物与环境之间气体交换、水分扩散中发挥重要作用. ...
Proteomics of terpenoid biosynthesis and secretion in trichomes of higher plant species
1
2016
... 植物一生固着于地面,因而更易遭受生物和非生物逆境胁迫.在长期进化过程中,植物从形态结构、生理生化等诸多方面演化形成一套完整的适应机制以抵御逆境胁迫.叶片作为地上部感知外界环境最敏感器官,其形态结构表现出与环境之间良好的适应协同性,如叶片表面蜡质可以抵御病原微生物和虫害以及其他生物胁迫和非生物胁迫[1~2],表皮腺体分泌物对生物胁迫和非生物胁迫[3],角质膜调节非气孔性蒸腾与体表温度[4]和防御病虫害叮咬[5~6]等.气孔是植物叶片特有的结构,在调节植物与环境之间气体交换、水分扩散中发挥重要作用. ...
A functional cutin matrix is required for plant protection against water loss
1
2011
... 植物一生固着于地面,因而更易遭受生物和非生物逆境胁迫.在长期进化过程中,植物从形态结构、生理生化等诸多方面演化形成一套完整的适应机制以抵御逆境胁迫.叶片作为地上部感知外界环境最敏感器官,其形态结构表现出与环境之间良好的适应协同性,如叶片表面蜡质可以抵御病原微生物和虫害以及其他生物胁迫和非生物胁迫[1~2],表皮腺体分泌物对生物胁迫和非生物胁迫[3],角质膜调节非气孔性蒸腾与体表温度[4]和防御病虫害叮咬[5~6]等.气孔是植物叶片特有的结构,在调节植物与环境之间气体交换、水分扩散中发挥重要作用. ...
Clytostoma callistegioides(Bignoniaceae) wax extract with activity on aphid settling
1
2010
... 植物一生固着于地面,因而更易遭受生物和非生物逆境胁迫.在长期进化过程中,植物从形态结构、生理生化等诸多方面演化形成一套完整的适应机制以抵御逆境胁迫.叶片作为地上部感知外界环境最敏感器官,其形态结构表现出与环境之间良好的适应协同性,如叶片表面蜡质可以抵御病原微生物和虫害以及其他生物胁迫和非生物胁迫[1~2],表皮腺体分泌物对生物胁迫和非生物胁迫[3],角质膜调节非气孔性蒸腾与体表温度[4]和防御病虫害叮咬[5~6]等.气孔是植物叶片特有的结构,在调节植物与环境之间气体交换、水分扩散中发挥重要作用. ...
Very-long-chain aldehydes promote in vitro prepenetration processes of Blumeria graminis in a dose- and chain length-dependent manner
1
2010
... 植物一生固着于地面,因而更易遭受生物和非生物逆境胁迫.在长期进化过程中,植物从形态结构、生理生化等诸多方面演化形成一套完整的适应机制以抵御逆境胁迫.叶片作为地上部感知外界环境最敏感器官,其形态结构表现出与环境之间良好的适应协同性,如叶片表面蜡质可以抵御病原微生物和虫害以及其他生物胁迫和非生物胁迫[1~2],表皮腺体分泌物对生物胁迫和非生物胁迫[3],角质膜调节非气孔性蒸腾与体表温度[4]和防御病虫害叮咬[5~6]等.气孔是植物叶片特有的结构,在调节植物与环境之间气体交换、水分扩散中发挥重要作用. ...
1
2015
... 干旱胁迫下植物叶片常通过调节气孔导度,增加气孔阻力以减少水分丧失.此外气孔密度、气孔大小和气孔形态结构等也与植物抗旱性密切相关[7].腺毛(腺体)是陆生植物地上部分表面广泛分布的特殊结构,它们具有合成、贮存和分泌次生代谢产物以及反射强光降低叶温、减少蒸腾等功能,在植物防御外界生物和非生物胁迫中起着重要作用.因此,了解叶片气孔、腺体等结构特征以及对逆境胁迫的响应是阐明植物抗逆机理的重要内容.甘草(Glycyrrhiza uralensis Fisch.)是豆科(Leguminosae)甘草属(Glycyrrhiza)多年生草本植物[8],多生长在干旱、半干旱荒漠草原、沙漠边缘和黄土丘陵地带.其根系发达,抗性强,是荒漠半荒漠地区改良土壤、保持水土、防风固沙的重要植物.目前,关于甘草抗旱性研究多集中在植株抗氧化系统[9~11]、种子萌发特性[12]、次生代谢[13~14]等方面,对干旱胁迫下甘草叶片形态结构研究较少.本试验采用盆栽控水模拟土壤干旱的方法,研究不同干旱胁迫程度对甘草幼苗叶片表面结构特征和光合作用的影响,旨在了解叶片表面结构对干旱逆境的响应特征,为阐明甘草抗旱机理提供重要的形态学和生理学依据. ...
1
2015
... 干旱胁迫下植物叶片常通过调节气孔导度,增加气孔阻力以减少水分丧失.此外气孔密度、气孔大小和气孔形态结构等也与植物抗旱性密切相关[7].腺毛(腺体)是陆生植物地上部分表面广泛分布的特殊结构,它们具有合成、贮存和分泌次生代谢产物以及反射强光降低叶温、减少蒸腾等功能,在植物防御外界生物和非生物胁迫中起着重要作用.因此,了解叶片气孔、腺体等结构特征以及对逆境胁迫的响应是阐明植物抗逆机理的重要内容.甘草(Glycyrrhiza uralensis Fisch.)是豆科(Leguminosae)甘草属(Glycyrrhiza)多年生草本植物[8],多生长在干旱、半干旱荒漠草原、沙漠边缘和黄土丘陵地带.其根系发达,抗性强,是荒漠半荒漠地区改良土壤、保持水土、防风固沙的重要植物.目前,关于甘草抗旱性研究多集中在植株抗氧化系统[9~11]、种子萌发特性[12]、次生代谢[13~14]等方面,对干旱胁迫下甘草叶片形态结构研究较少.本试验采用盆栽控水模拟土壤干旱的方法,研究不同干旱胁迫程度对甘草幼苗叶片表面结构特征和光合作用的影响,旨在了解叶片表面结构对干旱逆境的响应特征,为阐明甘草抗旱机理提供重要的形态学和生理学依据. ...
1
1998
... 干旱胁迫下植物叶片常通过调节气孔导度,增加气孔阻力以减少水分丧失.此外气孔密度、气孔大小和气孔形态结构等也与植物抗旱性密切相关[7].腺毛(腺体)是陆生植物地上部分表面广泛分布的特殊结构,它们具有合成、贮存和分泌次生代谢产物以及反射强光降低叶温、减少蒸腾等功能,在植物防御外界生物和非生物胁迫中起着重要作用.因此,了解叶片气孔、腺体等结构特征以及对逆境胁迫的响应是阐明植物抗逆机理的重要内容.甘草(Glycyrrhiza uralensis Fisch.)是豆科(Leguminosae)甘草属(Glycyrrhiza)多年生草本植物[8],多生长在干旱、半干旱荒漠草原、沙漠边缘和黄土丘陵地带.其根系发达,抗性强,是荒漠半荒漠地区改良土壤、保持水土、防风固沙的重要植物.目前,关于甘草抗旱性研究多集中在植株抗氧化系统[9~11]、种子萌发特性[12]、次生代谢[13~14]等方面,对干旱胁迫下甘草叶片形态结构研究较少.本试验采用盆栽控水模拟土壤干旱的方法,研究不同干旱胁迫程度对甘草幼苗叶片表面结构特征和光合作用的影响,旨在了解叶片表面结构对干旱逆境的响应特征,为阐明甘草抗旱机理提供重要的形态学和生理学依据. ...
1
1998
... 干旱胁迫下植物叶片常通过调节气孔导度,增加气孔阻力以减少水分丧失.此外气孔密度、气孔大小和气孔形态结构等也与植物抗旱性密切相关[7].腺毛(腺体)是陆生植物地上部分表面广泛分布的特殊结构,它们具有合成、贮存和分泌次生代谢产物以及反射强光降低叶温、减少蒸腾等功能,在植物防御外界生物和非生物胁迫中起着重要作用.因此,了解叶片气孔、腺体等结构特征以及对逆境胁迫的响应是阐明植物抗逆机理的重要内容.甘草(Glycyrrhiza uralensis Fisch.)是豆科(Leguminosae)甘草属(Glycyrrhiza)多年生草本植物[8],多生长在干旱、半干旱荒漠草原、沙漠边缘和黄土丘陵地带.其根系发达,抗性强,是荒漠半荒漠地区改良土壤、保持水土、防风固沙的重要植物.目前,关于甘草抗旱性研究多集中在植株抗氧化系统[9~11]、种子萌发特性[12]、次生代谢[13~14]等方面,对干旱胁迫下甘草叶片形态结构研究较少.本试验采用盆栽控水模拟土壤干旱的方法,研究不同干旱胁迫程度对甘草幼苗叶片表面结构特征和光合作用的影响,旨在了解叶片表面结构对干旱逆境的响应特征,为阐明甘草抗旱机理提供重要的形态学和生理学依据. ...
旱盐双重胁迫对乌拉尔甘草幼苗生理生化特性的影响
1
2018
... 干旱胁迫下植物叶片常通过调节气孔导度,增加气孔阻力以减少水分丧失.此外气孔密度、气孔大小和气孔形态结构等也与植物抗旱性密切相关[7].腺毛(腺体)是陆生植物地上部分表面广泛分布的特殊结构,它们具有合成、贮存和分泌次生代谢产物以及反射强光降低叶温、减少蒸腾等功能,在植物防御外界生物和非生物胁迫中起着重要作用.因此,了解叶片气孔、腺体等结构特征以及对逆境胁迫的响应是阐明植物抗逆机理的重要内容.甘草(Glycyrrhiza uralensis Fisch.)是豆科(Leguminosae)甘草属(Glycyrrhiza)多年生草本植物[8],多生长在干旱、半干旱荒漠草原、沙漠边缘和黄土丘陵地带.其根系发达,抗性强,是荒漠半荒漠地区改良土壤、保持水土、防风固沙的重要植物.目前,关于甘草抗旱性研究多集中在植株抗氧化系统[9~11]、种子萌发特性[12]、次生代谢[13~14]等方面,对干旱胁迫下甘草叶片形态结构研究较少.本试验采用盆栽控水模拟土壤干旱的方法,研究不同干旱胁迫程度对甘草幼苗叶片表面结构特征和光合作用的影响,旨在了解叶片表面结构对干旱逆境的响应特征,为阐明甘草抗旱机理提供重要的形态学和生理学依据. ...
旱盐双重胁迫对乌拉尔甘草幼苗生理生化特性的影响
1
2018
... 干旱胁迫下植物叶片常通过调节气孔导度,增加气孔阻力以减少水分丧失.此外气孔密度、气孔大小和气孔形态结构等也与植物抗旱性密切相关[7].腺毛(腺体)是陆生植物地上部分表面广泛分布的特殊结构,它们具有合成、贮存和分泌次生代谢产物以及反射强光降低叶温、减少蒸腾等功能,在植物防御外界生物和非生物胁迫中起着重要作用.因此,了解叶片气孔、腺体等结构特征以及对逆境胁迫的响应是阐明植物抗逆机理的重要内容.甘草(Glycyrrhiza uralensis Fisch.)是豆科(Leguminosae)甘草属(Glycyrrhiza)多年生草本植物[8],多生长在干旱、半干旱荒漠草原、沙漠边缘和黄土丘陵地带.其根系发达,抗性强,是荒漠半荒漠地区改良土壤、保持水土、防风固沙的重要植物.目前,关于甘草抗旱性研究多集中在植株抗氧化系统[9~11]、种子萌发特性[12]、次生代谢[13~14]等方面,对干旱胁迫下甘草叶片形态结构研究较少.本试验采用盆栽控水模拟土壤干旱的方法,研究不同干旱胁迫程度对甘草幼苗叶片表面结构特征和光合作用的影响,旨在了解叶片表面结构对干旱逆境的响应特征,为阐明甘草抗旱机理提供重要的形态学和生理学依据. ...
双重胁迫对乌拉尔甘草种子萌发及幼苗生理特性的影响
0
2018
双重胁迫对乌拉尔甘草种子萌发及幼苗生理特性的影响
0
2018
短小芽孢杆菌对盐胁迫下甘草生长及抗氧化系统的影响
1
2019
... 干旱胁迫下植物叶片常通过调节气孔导度,增加气孔阻力以减少水分丧失.此外气孔密度、气孔大小和气孔形态结构等也与植物抗旱性密切相关[7].腺毛(腺体)是陆生植物地上部分表面广泛分布的特殊结构,它们具有合成、贮存和分泌次生代谢产物以及反射强光降低叶温、减少蒸腾等功能,在植物防御外界生物和非生物胁迫中起着重要作用.因此,了解叶片气孔、腺体等结构特征以及对逆境胁迫的响应是阐明植物抗逆机理的重要内容.甘草(Glycyrrhiza uralensis Fisch.)是豆科(Leguminosae)甘草属(Glycyrrhiza)多年生草本植物[8],多生长在干旱、半干旱荒漠草原、沙漠边缘和黄土丘陵地带.其根系发达,抗性强,是荒漠半荒漠地区改良土壤、保持水土、防风固沙的重要植物.目前,关于甘草抗旱性研究多集中在植株抗氧化系统[9~11]、种子萌发特性[12]、次生代谢[13~14]等方面,对干旱胁迫下甘草叶片形态结构研究较少.本试验采用盆栽控水模拟土壤干旱的方法,研究不同干旱胁迫程度对甘草幼苗叶片表面结构特征和光合作用的影响,旨在了解叶片表面结构对干旱逆境的响应特征,为阐明甘草抗旱机理提供重要的形态学和生理学依据. ...
短小芽孢杆菌对盐胁迫下甘草生长及抗氧化系统的影响
1
2019
... 干旱胁迫下植物叶片常通过调节气孔导度,增加气孔阻力以减少水分丧失.此外气孔密度、气孔大小和气孔形态结构等也与植物抗旱性密切相关[7].腺毛(腺体)是陆生植物地上部分表面广泛分布的特殊结构,它们具有合成、贮存和分泌次生代谢产物以及反射强光降低叶温、减少蒸腾等功能,在植物防御外界生物和非生物胁迫中起着重要作用.因此,了解叶片气孔、腺体等结构特征以及对逆境胁迫的响应是阐明植物抗逆机理的重要内容.甘草(Glycyrrhiza uralensis Fisch.)是豆科(Leguminosae)甘草属(Glycyrrhiza)多年生草本植物[8],多生长在干旱、半干旱荒漠草原、沙漠边缘和黄土丘陵地带.其根系发达,抗性强,是荒漠半荒漠地区改良土壤、保持水土、防风固沙的重要植物.目前,关于甘草抗旱性研究多集中在植株抗氧化系统[9~11]、种子萌发特性[12]、次生代谢[13~14]等方面,对干旱胁迫下甘草叶片形态结构研究较少.本试验采用盆栽控水模拟土壤干旱的方法,研究不同干旱胁迫程度对甘草幼苗叶片表面结构特征和光合作用的影响,旨在了解叶片表面结构对干旱逆境的响应特征,为阐明甘草抗旱机理提供重要的形态学和生理学依据. ...
2种药用甘草种子对盐渍环境的萌发响应及其甘草酸含量变化
1
2018
... 干旱胁迫下植物叶片常通过调节气孔导度,增加气孔阻力以减少水分丧失.此外气孔密度、气孔大小和气孔形态结构等也与植物抗旱性密切相关[7].腺毛(腺体)是陆生植物地上部分表面广泛分布的特殊结构,它们具有合成、贮存和分泌次生代谢产物以及反射强光降低叶温、减少蒸腾等功能,在植物防御外界生物和非生物胁迫中起着重要作用.因此,了解叶片气孔、腺体等结构特征以及对逆境胁迫的响应是阐明植物抗逆机理的重要内容.甘草(Glycyrrhiza uralensis Fisch.)是豆科(Leguminosae)甘草属(Glycyrrhiza)多年生草本植物[8],多生长在干旱、半干旱荒漠草原、沙漠边缘和黄土丘陵地带.其根系发达,抗性强,是荒漠半荒漠地区改良土壤、保持水土、防风固沙的重要植物.目前,关于甘草抗旱性研究多集中在植株抗氧化系统[9~11]、种子萌发特性[12]、次生代谢[13~14]等方面,对干旱胁迫下甘草叶片形态结构研究较少.本试验采用盆栽控水模拟土壤干旱的方法,研究不同干旱胁迫程度对甘草幼苗叶片表面结构特征和光合作用的影响,旨在了解叶片表面结构对干旱逆境的响应特征,为阐明甘草抗旱机理提供重要的形态学和生理学依据. ...
2种药用甘草种子对盐渍环境的萌发响应及其甘草酸含量变化
1
2018
... 干旱胁迫下植物叶片常通过调节气孔导度,增加气孔阻力以减少水分丧失.此外气孔密度、气孔大小和气孔形态结构等也与植物抗旱性密切相关[7].腺毛(腺体)是陆生植物地上部分表面广泛分布的特殊结构,它们具有合成、贮存和分泌次生代谢产物以及反射强光降低叶温、减少蒸腾等功能,在植物防御外界生物和非生物胁迫中起着重要作用.因此,了解叶片气孔、腺体等结构特征以及对逆境胁迫的响应是阐明植物抗逆机理的重要内容.甘草(Glycyrrhiza uralensis Fisch.)是豆科(Leguminosae)甘草属(Glycyrrhiza)多年生草本植物[8],多生长在干旱、半干旱荒漠草原、沙漠边缘和黄土丘陵地带.其根系发达,抗性强,是荒漠半荒漠地区改良土壤、保持水土、防风固沙的重要植物.目前,关于甘草抗旱性研究多集中在植株抗氧化系统[9~11]、种子萌发特性[12]、次生代谢[13~14]等方面,对干旱胁迫下甘草叶片形态结构研究较少.本试验采用盆栽控水模拟土壤干旱的方法,研究不同干旱胁迫程度对甘草幼苗叶片表面结构特征和光合作用的影响,旨在了解叶片表面结构对干旱逆境的响应特征,为阐明甘草抗旱机理提供重要的形态学和生理学依据. ...
硅对盐胁迫下甘草非药用部位总黄酮、总皂苷积累动态的影响
1
2018
... 干旱胁迫下植物叶片常通过调节气孔导度,增加气孔阻力以减少水分丧失.此外气孔密度、气孔大小和气孔形态结构等也与植物抗旱性密切相关[7].腺毛(腺体)是陆生植物地上部分表面广泛分布的特殊结构,它们具有合成、贮存和分泌次生代谢产物以及反射强光降低叶温、减少蒸腾等功能,在植物防御外界生物和非生物胁迫中起着重要作用.因此,了解叶片气孔、腺体等结构特征以及对逆境胁迫的响应是阐明植物抗逆机理的重要内容.甘草(Glycyrrhiza uralensis Fisch.)是豆科(Leguminosae)甘草属(Glycyrrhiza)多年生草本植物[8],多生长在干旱、半干旱荒漠草原、沙漠边缘和黄土丘陵地带.其根系发达,抗性强,是荒漠半荒漠地区改良土壤、保持水土、防风固沙的重要植物.目前,关于甘草抗旱性研究多集中在植株抗氧化系统[9~11]、种子萌发特性[12]、次生代谢[13~14]等方面,对干旱胁迫下甘草叶片形态结构研究较少.本试验采用盆栽控水模拟土壤干旱的方法,研究不同干旱胁迫程度对甘草幼苗叶片表面结构特征和光合作用的影响,旨在了解叶片表面结构对干旱逆境的响应特征,为阐明甘草抗旱机理提供重要的形态学和生理学依据. ...
硅对盐胁迫下甘草非药用部位总黄酮、总皂苷积累动态的影响
1
2018
... 干旱胁迫下植物叶片常通过调节气孔导度,增加气孔阻力以减少水分丧失.此外气孔密度、气孔大小和气孔形态结构等也与植物抗旱性密切相关[7].腺毛(腺体)是陆生植物地上部分表面广泛分布的特殊结构,它们具有合成、贮存和分泌次生代谢产物以及反射强光降低叶温、减少蒸腾等功能,在植物防御外界生物和非生物胁迫中起着重要作用.因此,了解叶片气孔、腺体等结构特征以及对逆境胁迫的响应是阐明植物抗逆机理的重要内容.甘草(Glycyrrhiza uralensis Fisch.)是豆科(Leguminosae)甘草属(Glycyrrhiza)多年生草本植物[8],多生长在干旱、半干旱荒漠草原、沙漠边缘和黄土丘陵地带.其根系发达,抗性强,是荒漠半荒漠地区改良土壤、保持水土、防风固沙的重要植物.目前,关于甘草抗旱性研究多集中在植株抗氧化系统[9~11]、种子萌发特性[12]、次生代谢[13~14]等方面,对干旱胁迫下甘草叶片形态结构研究较少.本试验采用盆栽控水模拟土壤干旱的方法,研究不同干旱胁迫程度对甘草幼苗叶片表面结构特征和光合作用的影响,旨在了解叶片表面结构对干旱逆境的响应特征,为阐明甘草抗旱机理提供重要的形态学和生理学依据. ...
盐生植物胀果甘草和光果甘草对UV-B以及盐胁迫的不同响应
1
2018
... 干旱胁迫下植物叶片常通过调节气孔导度,增加气孔阻力以减少水分丧失.此外气孔密度、气孔大小和气孔形态结构等也与植物抗旱性密切相关[7].腺毛(腺体)是陆生植物地上部分表面广泛分布的特殊结构,它们具有合成、贮存和分泌次生代谢产物以及反射强光降低叶温、减少蒸腾等功能,在植物防御外界生物和非生物胁迫中起着重要作用.因此,了解叶片气孔、腺体等结构特征以及对逆境胁迫的响应是阐明植物抗逆机理的重要内容.甘草(Glycyrrhiza uralensis Fisch.)是豆科(Leguminosae)甘草属(Glycyrrhiza)多年生草本植物[8],多生长在干旱、半干旱荒漠草原、沙漠边缘和黄土丘陵地带.其根系发达,抗性强,是荒漠半荒漠地区改良土壤、保持水土、防风固沙的重要植物.目前,关于甘草抗旱性研究多集中在植株抗氧化系统[9~11]、种子萌发特性[12]、次生代谢[13~14]等方面,对干旱胁迫下甘草叶片形态结构研究较少.本试验采用盆栽控水模拟土壤干旱的方法,研究不同干旱胁迫程度对甘草幼苗叶片表面结构特征和光合作用的影响,旨在了解叶片表面结构对干旱逆境的响应特征,为阐明甘草抗旱机理提供重要的形态学和生理学依据. ...
盐生植物胀果甘草和光果甘草对UV-B以及盐胁迫的不同响应
1
2018
... 干旱胁迫下植物叶片常通过调节气孔导度,增加气孔阻力以减少水分丧失.此外气孔密度、气孔大小和气孔形态结构等也与植物抗旱性密切相关[7].腺毛(腺体)是陆生植物地上部分表面广泛分布的特殊结构,它们具有合成、贮存和分泌次生代谢产物以及反射强光降低叶温、减少蒸腾等功能,在植物防御外界生物和非生物胁迫中起着重要作用.因此,了解叶片气孔、腺体等结构特征以及对逆境胁迫的响应是阐明植物抗逆机理的重要内容.甘草(Glycyrrhiza uralensis Fisch.)是豆科(Leguminosae)甘草属(Glycyrrhiza)多年生草本植物[8],多生长在干旱、半干旱荒漠草原、沙漠边缘和黄土丘陵地带.其根系发达,抗性强,是荒漠半荒漠地区改良土壤、保持水土、防风固沙的重要植物.目前,关于甘草抗旱性研究多集中在植株抗氧化系统[9~11]、种子萌发特性[12]、次生代谢[13~14]等方面,对干旱胁迫下甘草叶片形态结构研究较少.本试验采用盆栽控水模拟土壤干旱的方法,研究不同干旱胁迫程度对甘草幼苗叶片表面结构特征和光合作用的影响,旨在了解叶片表面结构对干旱逆境的响应特征,为阐明甘草抗旱机理提供重要的形态学和生理学依据. ...
植物叶片扫描电镜样品制备
1
1988
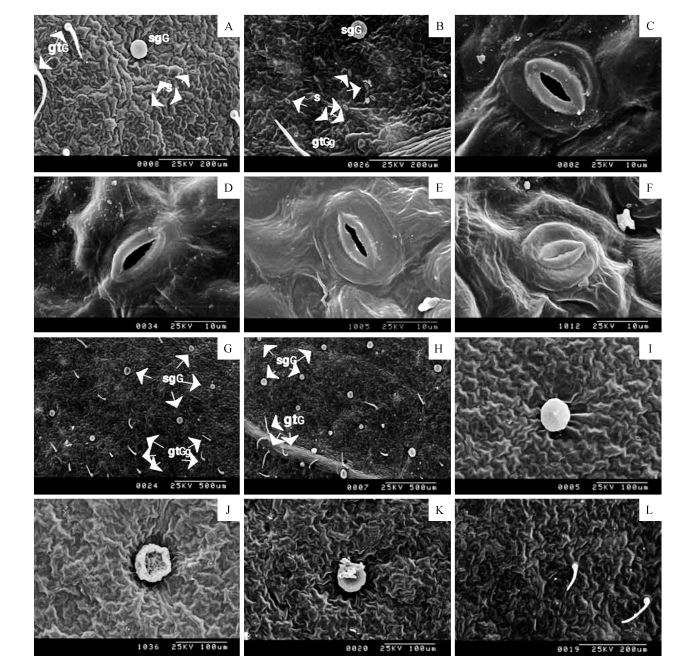
... 参照李柏年等[16]方法略有改动.取每个处理甘草植株顶端新叶、中部成熟叶和基部衰老叶,避开主脉,用刀片分别切取8 mm×5 mm的叶肉组织,2.5%戊二醛固定2 h,抽气下沉,不同浓度乙醇梯度脱水,临界点干燥及喷金处理,扫描电镜观察并照相.每个处理选取10个视野,统计气孔密度、气孔开张比并测量气孔开张度.气孔开张度为每个视野随机选取10个开张气孔,测量气孔孔径宽度并计算平均值;气孔开张比为每个视野中开张的气孔数占总气孔数的百分比. ...
植物叶片扫描电镜样品制备
1
1988
... 参照李柏年等[16]方法略有改动.取每个处理甘草植株顶端新叶、中部成熟叶和基部衰老叶,避开主脉,用刀片分别切取8 mm×5 mm的叶肉组织,2.5%戊二醛固定2 h,抽气下沉,不同浓度乙醇梯度脱水,临界点干燥及喷金处理,扫描电镜观察并照相.每个处理选取10个视野,统计气孔密度、气孔开张比并测量气孔开张度.气孔开张度为每个视野随机选取10个开张气孔,测量气孔孔径宽度并计算平均值;气孔开张比为每个视野中开张的气孔数占总气孔数的百分比. ...
气孔特征与叶片功能性状之间关联性沿海拔梯度的变化规律——以长白山为例
1
2016
... 叶片是植物进行光合作用的主要器官,气孔是叶表面特有的形态结构,植物通过气孔与外界环境进行水分和气体交换,气孔开度控制着植物的光合作用以及蒸腾作用[17~18],从而影响叶片水势.因此,气孔的形态建成、分布密度以及开放程度等均直接影响植物光合作用以及蒸腾作用.大量研究均证实,气孔在叶表面分布不均,主要分布在下表面[19].本研究同样观察到,甘草叶片气孔在下表面分布密度远高于上表面.正常供水条件下,不同发育程度叶片表皮气孔密度成熟叶>新叶>衰老叶,以及气孔开张度也表现为成熟叶>新叶>衰老叶.这可能是随着分生组织细胞成熟,最终分裂为气孔,因而随着叶片成熟,单位面积上气孔数量增多[20],然而随着叶片衰老,气孔也随着发生PCD,继而气孔密度减小. ...
气孔特征与叶片功能性状之间关联性沿海拔梯度的变化规律——以长白山为例
1
2016
... 叶片是植物进行光合作用的主要器官,气孔是叶表面特有的形态结构,植物通过气孔与外界环境进行水分和气体交换,气孔开度控制着植物的光合作用以及蒸腾作用[17~18],从而影响叶片水势.因此,气孔的形态建成、分布密度以及开放程度等均直接影响植物光合作用以及蒸腾作用.大量研究均证实,气孔在叶表面分布不均,主要分布在下表面[19].本研究同样观察到,甘草叶片气孔在下表面分布密度远高于上表面.正常供水条件下,不同发育程度叶片表皮气孔密度成熟叶>新叶>衰老叶,以及气孔开张度也表现为成熟叶>新叶>衰老叶.这可能是随着分生组织细胞成熟,最终分裂为气孔,因而随着叶片成熟,单位面积上气孔数量增多[20],然而随着叶片衰老,气孔也随着发生PCD,继而气孔密度减小. ...
The role of stomata in sensing and driving environmental change
1
2003
... 叶片是植物进行光合作用的主要器官,气孔是叶表面特有的形态结构,植物通过气孔与外界环境进行水分和气体交换,气孔开度控制着植物的光合作用以及蒸腾作用[17~18],从而影响叶片水势.因此,气孔的形态建成、分布密度以及开放程度等均直接影响植物光合作用以及蒸腾作用.大量研究均证实,气孔在叶表面分布不均,主要分布在下表面[19].本研究同样观察到,甘草叶片气孔在下表面分布密度远高于上表面.正常供水条件下,不同发育程度叶片表皮气孔密度成熟叶>新叶>衰老叶,以及气孔开张度也表现为成熟叶>新叶>衰老叶.这可能是随着分生组织细胞成熟,最终分裂为气孔,因而随着叶片成熟,单位面积上气孔数量增多[20],然而随着叶片衰老,气孔也随着发生PCD,继而气孔密度减小. ...
8种绿化树种光合特性及叶片解剖结构比较
1
2019
... 叶片是植物进行光合作用的主要器官,气孔是叶表面特有的形态结构,植物通过气孔与外界环境进行水分和气体交换,气孔开度控制着植物的光合作用以及蒸腾作用[17~18],从而影响叶片水势.因此,气孔的形态建成、分布密度以及开放程度等均直接影响植物光合作用以及蒸腾作用.大量研究均证实,气孔在叶表面分布不均,主要分布在下表面[19].本研究同样观察到,甘草叶片气孔在下表面分布密度远高于上表面.正常供水条件下,不同发育程度叶片表皮气孔密度成熟叶>新叶>衰老叶,以及气孔开张度也表现为成熟叶>新叶>衰老叶.这可能是随着分生组织细胞成熟,最终分裂为气孔,因而随着叶片成熟,单位面积上气孔数量增多[20],然而随着叶片衰老,气孔也随着发生PCD,继而气孔密度减小. ...
8种绿化树种光合特性及叶片解剖结构比较
1
2019
... 叶片是植物进行光合作用的主要器官,气孔是叶表面特有的形态结构,植物通过气孔与外界环境进行水分和气体交换,气孔开度控制着植物的光合作用以及蒸腾作用[17~18],从而影响叶片水势.因此,气孔的形态建成、分布密度以及开放程度等均直接影响植物光合作用以及蒸腾作用.大量研究均证实,气孔在叶表面分布不均,主要分布在下表面[19].本研究同样观察到,甘草叶片气孔在下表面分布密度远高于上表面.正常供水条件下,不同发育程度叶片表皮气孔密度成熟叶>新叶>衰老叶,以及气孔开张度也表现为成熟叶>新叶>衰老叶.这可能是随着分生组织细胞成熟,最终分裂为气孔,因而随着叶片成熟,单位面积上气孔数量增多[20],然而随着叶片衰老,气孔也随着发生PCD,继而气孔密度减小. ...
Oriented asymmetric divisions that generate the stomatal spacing pattern in Arabidopsis are disrupted by the too many mouths mutation
1
2000
... 叶片是植物进行光合作用的主要器官,气孔是叶表面特有的形态结构,植物通过气孔与外界环境进行水分和气体交换,气孔开度控制着植物的光合作用以及蒸腾作用[17~18],从而影响叶片水势.因此,气孔的形态建成、分布密度以及开放程度等均直接影响植物光合作用以及蒸腾作用.大量研究均证实,气孔在叶表面分布不均,主要分布在下表面[19].本研究同样观察到,甘草叶片气孔在下表面分布密度远高于上表面.正常供水条件下,不同发育程度叶片表皮气孔密度成熟叶>新叶>衰老叶,以及气孔开张度也表现为成熟叶>新叶>衰老叶.这可能是随着分生组织细胞成熟,最终分裂为气孔,因而随着叶片成熟,单位面积上气孔数量增多[20],然而随着叶片衰老,气孔也随着发生PCD,继而气孔密度减小. ...
不同甘蔗品种叶片气孔对水分胁迫的响应
1
2014
... 植物抵御干旱胁迫主动适应环境的一个重要调节因素就是气孔调节[21],干旱胁迫会诱导植物的气孔出现微形态的变化,如气孔密度增大[22]、气孔开度减小[23~24]、气孔下陷、气孔器长度和宽度显著减小等[25],以使植物抗性提高.本试验结果显示,随着干旱胁迫程度的加剧,叶片总气孔密度、气孔开张比呈先增大后减小的趋势;而气孔开张宽度呈逐渐减小的趋势,可能是气孔密度的增加有助于植物更好地控制呼吸作用,调节体内H2O和CO2的进出,同时脱落酸(ABA)触发保卫细胞释放K+,保卫细胞渗透压和体积的降低导致气孔开张宽度逐渐减小,甚至关闭.与张晓艳等[26]研究发现气孔密度随干早程度的加剧呈先增加后减小结果一致,说明干旱胁迫程度加速了气孔细胞的程序性死亡所造成的[27],与赵殊丽[28]、齐红岩等[29]、高彦萍等[30]的研究气孔密度逐渐增大结果不同,这可能是甘草叶片具有表皮毛、腺体等适应干旱的特征,对干旱逆境胁迫具有交叉适应性,既能提高植株的抗旱能力,又能降低其在干旱环境下所受到的伤害. ...
不同甘蔗品种叶片气孔对水分胁迫的响应
1
2014
... 植物抵御干旱胁迫主动适应环境的一个重要调节因素就是气孔调节[21],干旱胁迫会诱导植物的气孔出现微形态的变化,如气孔密度增大[22]、气孔开度减小[23~24]、气孔下陷、气孔器长度和宽度显著减小等[25],以使植物抗性提高.本试验结果显示,随着干旱胁迫程度的加剧,叶片总气孔密度、气孔开张比呈先增大后减小的趋势;而气孔开张宽度呈逐渐减小的趋势,可能是气孔密度的增加有助于植物更好地控制呼吸作用,调节体内H2O和CO2的进出,同时脱落酸(ABA)触发保卫细胞释放K+,保卫细胞渗透压和体积的降低导致气孔开张宽度逐渐减小,甚至关闭.与张晓艳等[26]研究发现气孔密度随干早程度的加剧呈先增加后减小结果一致,说明干旱胁迫程度加速了气孔细胞的程序性死亡所造成的[27],与赵殊丽[28]、齐红岩等[29]、高彦萍等[30]的研究气孔密度逐渐增大结果不同,这可能是甘草叶片具有表皮毛、腺体等适应干旱的特征,对干旱逆境胁迫具有交叉适应性,既能提高植株的抗旱能力,又能降低其在干旱环境下所受到的伤害. ...
Photosynthetic characteristics during on togenesis of leaves.7.Stomata density and sizes
1
1982
... 植物抵御干旱胁迫主动适应环境的一个重要调节因素就是气孔调节[21],干旱胁迫会诱导植物的气孔出现微形态的变化,如气孔密度增大[22]、气孔开度减小[23~24]、气孔下陷、气孔器长度和宽度显著减小等[25],以使植物抗性提高.本试验结果显示,随着干旱胁迫程度的加剧,叶片总气孔密度、气孔开张比呈先增大后减小的趋势;而气孔开张宽度呈逐渐减小的趋势,可能是气孔密度的增加有助于植物更好地控制呼吸作用,调节体内H2O和CO2的进出,同时脱落酸(ABA)触发保卫细胞释放K+,保卫细胞渗透压和体积的降低导致气孔开张宽度逐渐减小,甚至关闭.与张晓艳等[26]研究发现气孔密度随干早程度的加剧呈先增加后减小结果一致,说明干旱胁迫程度加速了气孔细胞的程序性死亡所造成的[27],与赵殊丽[28]、齐红岩等[29]、高彦萍等[30]的研究气孔密度逐渐增大结果不同,这可能是甘草叶片具有表皮毛、腺体等适应干旱的特征,对干旱逆境胁迫具有交叉适应性,既能提高植株的抗旱能力,又能降低其在干旱环境下所受到的伤害. ...
干旱胁迫下水杨酸对银沙槐子叶表皮气孔开度的影响
1
2014
... 植物抵御干旱胁迫主动适应环境的一个重要调节因素就是气孔调节[21],干旱胁迫会诱导植物的气孔出现微形态的变化,如气孔密度增大[22]、气孔开度减小[23~24]、气孔下陷、气孔器长度和宽度显著减小等[25],以使植物抗性提高.本试验结果显示,随着干旱胁迫程度的加剧,叶片总气孔密度、气孔开张比呈先增大后减小的趋势;而气孔开张宽度呈逐渐减小的趋势,可能是气孔密度的增加有助于植物更好地控制呼吸作用,调节体内H2O和CO2的进出,同时脱落酸(ABA)触发保卫细胞释放K+,保卫细胞渗透压和体积的降低导致气孔开张宽度逐渐减小,甚至关闭.与张晓艳等[26]研究发现气孔密度随干早程度的加剧呈先增加后减小结果一致,说明干旱胁迫程度加速了气孔细胞的程序性死亡所造成的[27],与赵殊丽[28]、齐红岩等[29]、高彦萍等[30]的研究气孔密度逐渐增大结果不同,这可能是甘草叶片具有表皮毛、腺体等适应干旱的特征,对干旱逆境胁迫具有交叉适应性,既能提高植株的抗旱能力,又能降低其在干旱环境下所受到的伤害. ...
干旱胁迫下水杨酸对银沙槐子叶表皮气孔开度的影响
1
2014
... 植物抵御干旱胁迫主动适应环境的一个重要调节因素就是气孔调节[21],干旱胁迫会诱导植物的气孔出现微形态的变化,如气孔密度增大[22]、气孔开度减小[23~24]、气孔下陷、气孔器长度和宽度显著减小等[25],以使植物抗性提高.本试验结果显示,随着干旱胁迫程度的加剧,叶片总气孔密度、气孔开张比呈先增大后减小的趋势;而气孔开张宽度呈逐渐减小的趋势,可能是气孔密度的增加有助于植物更好地控制呼吸作用,调节体内H2O和CO2的进出,同时脱落酸(ABA)触发保卫细胞释放K+,保卫细胞渗透压和体积的降低导致气孔开张宽度逐渐减小,甚至关闭.与张晓艳等[26]研究发现气孔密度随干早程度的加剧呈先增加后减小结果一致,说明干旱胁迫程度加速了气孔细胞的程序性死亡所造成的[27],与赵殊丽[28]、齐红岩等[29]、高彦萍等[30]的研究气孔密度逐渐增大结果不同,这可能是甘草叶片具有表皮毛、腺体等适应干旱的特征,对干旱逆境胁迫具有交叉适应性,既能提高植株的抗旱能力,又能降低其在干旱环境下所受到的伤害. ...
水分胁迫对短枝型果树光合作用的非气孔限制
1
2003
... 植物抵御干旱胁迫主动适应环境的一个重要调节因素就是气孔调节[21],干旱胁迫会诱导植物的气孔出现微形态的变化,如气孔密度增大[22]、气孔开度减小[23~24]、气孔下陷、气孔器长度和宽度显著减小等[25],以使植物抗性提高.本试验结果显示,随着干旱胁迫程度的加剧,叶片总气孔密度、气孔开张比呈先增大后减小的趋势;而气孔开张宽度呈逐渐减小的趋势,可能是气孔密度的增加有助于植物更好地控制呼吸作用,调节体内H2O和CO2的进出,同时脱落酸(ABA)触发保卫细胞释放K+,保卫细胞渗透压和体积的降低导致气孔开张宽度逐渐减小,甚至关闭.与张晓艳等[26]研究发现气孔密度随干早程度的加剧呈先增加后减小结果一致,说明干旱胁迫程度加速了气孔细胞的程序性死亡所造成的[27],与赵殊丽[28]、齐红岩等[29]、高彦萍等[30]的研究气孔密度逐渐增大结果不同,这可能是甘草叶片具有表皮毛、腺体等适应干旱的特征,对干旱逆境胁迫具有交叉适应性,既能提高植株的抗旱能力,又能降低其在干旱环境下所受到的伤害. ...
水分胁迫对短枝型果树光合作用的非气孔限制
1
2003
... 植物抵御干旱胁迫主动适应环境的一个重要调节因素就是气孔调节[21],干旱胁迫会诱导植物的气孔出现微形态的变化,如气孔密度增大[22]、气孔开度减小[23~24]、气孔下陷、气孔器长度和宽度显著减小等[25],以使植物抗性提高.本试验结果显示,随着干旱胁迫程度的加剧,叶片总气孔密度、气孔开张比呈先增大后减小的趋势;而气孔开张宽度呈逐渐减小的趋势,可能是气孔密度的增加有助于植物更好地控制呼吸作用,调节体内H2O和CO2的进出,同时脱落酸(ABA)触发保卫细胞释放K+,保卫细胞渗透压和体积的降低导致气孔开张宽度逐渐减小,甚至关闭.与张晓艳等[26]研究发现气孔密度随干早程度的加剧呈先增加后减小结果一致,说明干旱胁迫程度加速了气孔细胞的程序性死亡所造成的[27],与赵殊丽[28]、齐红岩等[29]、高彦萍等[30]的研究气孔密度逐渐增大结果不同,这可能是甘草叶片具有表皮毛、腺体等适应干旱的特征,对干旱逆境胁迫具有交叉适应性,既能提高植株的抗旱能力,又能降低其在干旱环境下所受到的伤害. ...
不同土壤水分含量对玉米气孔发育过程和蒸腾耗水量的影响
1
2011
... 植物抵御干旱胁迫主动适应环境的一个重要调节因素就是气孔调节[21],干旱胁迫会诱导植物的气孔出现微形态的变化,如气孔密度增大[22]、气孔开度减小[23~24]、气孔下陷、气孔器长度和宽度显著减小等[25],以使植物抗性提高.本试验结果显示,随着干旱胁迫程度的加剧,叶片总气孔密度、气孔开张比呈先增大后减小的趋势;而气孔开张宽度呈逐渐减小的趋势,可能是气孔密度的增加有助于植物更好地控制呼吸作用,调节体内H2O和CO2的进出,同时脱落酸(ABA)触发保卫细胞释放K+,保卫细胞渗透压和体积的降低导致气孔开张宽度逐渐减小,甚至关闭.与张晓艳等[26]研究发现气孔密度随干早程度的加剧呈先增加后减小结果一致,说明干旱胁迫程度加速了气孔细胞的程序性死亡所造成的[27],与赵殊丽[28]、齐红岩等[29]、高彦萍等[30]的研究气孔密度逐渐增大结果不同,这可能是甘草叶片具有表皮毛、腺体等适应干旱的特征,对干旱逆境胁迫具有交叉适应性,既能提高植株的抗旱能力,又能降低其在干旱环境下所受到的伤害. ...
不同土壤水分含量对玉米气孔发育过程和蒸腾耗水量的影响
1
2011
... 植物抵御干旱胁迫主动适应环境的一个重要调节因素就是气孔调节[21],干旱胁迫会诱导植物的气孔出现微形态的变化,如气孔密度增大[22]、气孔开度减小[23~24]、气孔下陷、气孔器长度和宽度显著减小等[25],以使植物抗性提高.本试验结果显示,随着干旱胁迫程度的加剧,叶片总气孔密度、气孔开张比呈先增大后减小的趋势;而气孔开张宽度呈逐渐减小的趋势,可能是气孔密度的增加有助于植物更好地控制呼吸作用,调节体内H2O和CO2的进出,同时脱落酸(ABA)触发保卫细胞释放K+,保卫细胞渗透压和体积的降低导致气孔开张宽度逐渐减小,甚至关闭.与张晓艳等[26]研究发现气孔密度随干早程度的加剧呈先增加后减小结果一致,说明干旱胁迫程度加速了气孔细胞的程序性死亡所造成的[27],与赵殊丽[28]、齐红岩等[29]、高彦萍等[30]的研究气孔密度逐渐增大结果不同,这可能是甘草叶片具有表皮毛、腺体等适应干旱的特征,对干旱逆境胁迫具有交叉适应性,既能提高植株的抗旱能力,又能降低其在干旱环境下所受到的伤害. ...
土壤水分和种植密度对春小麦叶片气孔的影响
1
2003
... 植物抵御干旱胁迫主动适应环境的一个重要调节因素就是气孔调节[21],干旱胁迫会诱导植物的气孔出现微形态的变化,如气孔密度增大[22]、气孔开度减小[23~24]、气孔下陷、气孔器长度和宽度显著减小等[25],以使植物抗性提高.本试验结果显示,随着干旱胁迫程度的加剧,叶片总气孔密度、气孔开张比呈先增大后减小的趋势;而气孔开张宽度呈逐渐减小的趋势,可能是气孔密度的增加有助于植物更好地控制呼吸作用,调节体内H2O和CO2的进出,同时脱落酸(ABA)触发保卫细胞释放K+,保卫细胞渗透压和体积的降低导致气孔开张宽度逐渐减小,甚至关闭.与张晓艳等[26]研究发现气孔密度随干早程度的加剧呈先增加后减小结果一致,说明干旱胁迫程度加速了气孔细胞的程序性死亡所造成的[27],与赵殊丽[28]、齐红岩等[29]、高彦萍等[30]的研究气孔密度逐渐增大结果不同,这可能是甘草叶片具有表皮毛、腺体等适应干旱的特征,对干旱逆境胁迫具有交叉适应性,既能提高植株的抗旱能力,又能降低其在干旱环境下所受到的伤害. ...
土壤水分和种植密度对春小麦叶片气孔的影响
1
2003
... 植物抵御干旱胁迫主动适应环境的一个重要调节因素就是气孔调节[21],干旱胁迫会诱导植物的气孔出现微形态的变化,如气孔密度增大[22]、气孔开度减小[23~24]、气孔下陷、气孔器长度和宽度显著减小等[25],以使植物抗性提高.本试验结果显示,随着干旱胁迫程度的加剧,叶片总气孔密度、气孔开张比呈先增大后减小的趋势;而气孔开张宽度呈逐渐减小的趋势,可能是气孔密度的增加有助于植物更好地控制呼吸作用,调节体内H2O和CO2的进出,同时脱落酸(ABA)触发保卫细胞释放K+,保卫细胞渗透压和体积的降低导致气孔开张宽度逐渐减小,甚至关闭.与张晓艳等[26]研究发现气孔密度随干早程度的加剧呈先增加后减小结果一致,说明干旱胁迫程度加速了气孔细胞的程序性死亡所造成的[27],与赵殊丽[28]、齐红岩等[29]、高彦萍等[30]的研究气孔密度逐渐增大结果不同,这可能是甘草叶片具有表皮毛、腺体等适应干旱的特征,对干旱逆境胁迫具有交叉适应性,既能提高植株的抗旱能力,又能降低其在干旱环境下所受到的伤害. ...
干旱和CO2浓度升高对干旱区春小麦气孔密度及分布的影响
1
2001
... 植物抵御干旱胁迫主动适应环境的一个重要调节因素就是气孔调节[21],干旱胁迫会诱导植物的气孔出现微形态的变化,如气孔密度增大[22]、气孔开度减小[23~24]、气孔下陷、气孔器长度和宽度显著减小等[25],以使植物抗性提高.本试验结果显示,随着干旱胁迫程度的加剧,叶片总气孔密度、气孔开张比呈先增大后减小的趋势;而气孔开张宽度呈逐渐减小的趋势,可能是气孔密度的增加有助于植物更好地控制呼吸作用,调节体内H2O和CO2的进出,同时脱落酸(ABA)触发保卫细胞释放K+,保卫细胞渗透压和体积的降低导致气孔开张宽度逐渐减小,甚至关闭.与张晓艳等[26]研究发现气孔密度随干早程度的加剧呈先增加后减小结果一致,说明干旱胁迫程度加速了气孔细胞的程序性死亡所造成的[27],与赵殊丽[28]、齐红岩等[29]、高彦萍等[30]的研究气孔密度逐渐增大结果不同,这可能是甘草叶片具有表皮毛、腺体等适应干旱的特征,对干旱逆境胁迫具有交叉适应性,既能提高植株的抗旱能力,又能降低其在干旱环境下所受到的伤害. ...
干旱和CO2浓度升高对干旱区春小麦气孔密度及分布的影响
1
2001
... 植物抵御干旱胁迫主动适应环境的一个重要调节因素就是气孔调节[21],干旱胁迫会诱导植物的气孔出现微形态的变化,如气孔密度增大[22]、气孔开度减小[23~24]、气孔下陷、气孔器长度和宽度显著减小等[25],以使植物抗性提高.本试验结果显示,随着干旱胁迫程度的加剧,叶片总气孔密度、气孔开张比呈先增大后减小的趋势;而气孔开张宽度呈逐渐减小的趋势,可能是气孔密度的增加有助于植物更好地控制呼吸作用,调节体内H2O和CO2的进出,同时脱落酸(ABA)触发保卫细胞释放K+,保卫细胞渗透压和体积的降低导致气孔开张宽度逐渐减小,甚至关闭.与张晓艳等[26]研究发现气孔密度随干早程度的加剧呈先增加后减小结果一致,说明干旱胁迫程度加速了气孔细胞的程序性死亡所造成的[27],与赵殊丽[28]、齐红岩等[29]、高彦萍等[30]的研究气孔密度逐渐增大结果不同,这可能是甘草叶片具有表皮毛、腺体等适应干旱的特征,对干旱逆境胁迫具有交叉适应性,既能提高植株的抗旱能力,又能降低其在干旱环境下所受到的伤害. ...
水分胁迫对水稻剑叶气孔特性的影响
1
2010
... 植物抵御干旱胁迫主动适应环境的一个重要调节因素就是气孔调节[21],干旱胁迫会诱导植物的气孔出现微形态的变化,如气孔密度增大[22]、气孔开度减小[23~24]、气孔下陷、气孔器长度和宽度显著减小等[25],以使植物抗性提高.本试验结果显示,随着干旱胁迫程度的加剧,叶片总气孔密度、气孔开张比呈先增大后减小的趋势;而气孔开张宽度呈逐渐减小的趋势,可能是气孔密度的增加有助于植物更好地控制呼吸作用,调节体内H2O和CO2的进出,同时脱落酸(ABA)触发保卫细胞释放K+,保卫细胞渗透压和体积的降低导致气孔开张宽度逐渐减小,甚至关闭.与张晓艳等[26]研究发现气孔密度随干早程度的加剧呈先增加后减小结果一致,说明干旱胁迫程度加速了气孔细胞的程序性死亡所造成的[27],与赵殊丽[28]、齐红岩等[29]、高彦萍等[30]的研究气孔密度逐渐增大结果不同,这可能是甘草叶片具有表皮毛、腺体等适应干旱的特征,对干旱逆境胁迫具有交叉适应性,既能提高植株的抗旱能力,又能降低其在干旱环境下所受到的伤害. ...
水分胁迫对水稻剑叶气孔特性的影响
1
2010
... 植物抵御干旱胁迫主动适应环境的一个重要调节因素就是气孔调节[21],干旱胁迫会诱导植物的气孔出现微形态的变化,如气孔密度增大[22]、气孔开度减小[23~24]、气孔下陷、气孔器长度和宽度显著减小等[25],以使植物抗性提高.本试验结果显示,随着干旱胁迫程度的加剧,叶片总气孔密度、气孔开张比呈先增大后减小的趋势;而气孔开张宽度呈逐渐减小的趋势,可能是气孔密度的增加有助于植物更好地控制呼吸作用,调节体内H2O和CO2的进出,同时脱落酸(ABA)触发保卫细胞释放K+,保卫细胞渗透压和体积的降低导致气孔开张宽度逐渐减小,甚至关闭.与张晓艳等[26]研究发现气孔密度随干早程度的加剧呈先增加后减小结果一致,说明干旱胁迫程度加速了气孔细胞的程序性死亡所造成的[27],与赵殊丽[28]、齐红岩等[29]、高彦萍等[30]的研究气孔密度逐渐增大结果不同,这可能是甘草叶片具有表皮毛、腺体等适应干旱的特征,对干旱逆境胁迫具有交叉适应性,既能提高植株的抗旱能力,又能降低其在干旱环境下所受到的伤害. ...
水分亏缺对番茄叶片气孔特性及叶绿体超微结构的影响
1
2009
... 植物抵御干旱胁迫主动适应环境的一个重要调节因素就是气孔调节[21],干旱胁迫会诱导植物的气孔出现微形态的变化,如气孔密度增大[22]、气孔开度减小[23~24]、气孔下陷、气孔器长度和宽度显著减小等[25],以使植物抗性提高.本试验结果显示,随着干旱胁迫程度的加剧,叶片总气孔密度、气孔开张比呈先增大后减小的趋势;而气孔开张宽度呈逐渐减小的趋势,可能是气孔密度的增加有助于植物更好地控制呼吸作用,调节体内H2O和CO2的进出,同时脱落酸(ABA)触发保卫细胞释放K+,保卫细胞渗透压和体积的降低导致气孔开张宽度逐渐减小,甚至关闭.与张晓艳等[26]研究发现气孔密度随干早程度的加剧呈先增加后减小结果一致,说明干旱胁迫程度加速了气孔细胞的程序性死亡所造成的[27],与赵殊丽[28]、齐红岩等[29]、高彦萍等[30]的研究气孔密度逐渐增大结果不同,这可能是甘草叶片具有表皮毛、腺体等适应干旱的特征,对干旱逆境胁迫具有交叉适应性,既能提高植株的抗旱能力,又能降低其在干旱环境下所受到的伤害. ...
水分亏缺对番茄叶片气孔特性及叶绿体超微结构的影响
1
2009
... 植物抵御干旱胁迫主动适应环境的一个重要调节因素就是气孔调节[21],干旱胁迫会诱导植物的气孔出现微形态的变化,如气孔密度增大[22]、气孔开度减小[23~24]、气孔下陷、气孔器长度和宽度显著减小等[25],以使植物抗性提高.本试验结果显示,随着干旱胁迫程度的加剧,叶片总气孔密度、气孔开张比呈先增大后减小的趋势;而气孔开张宽度呈逐渐减小的趋势,可能是气孔密度的增加有助于植物更好地控制呼吸作用,调节体内H2O和CO2的进出,同时脱落酸(ABA)触发保卫细胞释放K+,保卫细胞渗透压和体积的降低导致气孔开张宽度逐渐减小,甚至关闭.与张晓艳等[26]研究发现气孔密度随干早程度的加剧呈先增加后减小结果一致,说明干旱胁迫程度加速了气孔细胞的程序性死亡所造成的[27],与赵殊丽[28]、齐红岩等[29]、高彦萍等[30]的研究气孔密度逐渐增大结果不同,这可能是甘草叶片具有表皮毛、腺体等适应干旱的特征,对干旱逆境胁迫具有交叉适应性,既能提高植株的抗旱能力,又能降低其在干旱环境下所受到的伤害. ...
水分胁迫下不同抗旱类型大豆叶片气孔特性变化研究
1
2007
... 植物抵御干旱胁迫主动适应环境的一个重要调节因素就是气孔调节[21],干旱胁迫会诱导植物的气孔出现微形态的变化,如气孔密度增大[22]、气孔开度减小[23~24]、气孔下陷、气孔器长度和宽度显著减小等[25],以使植物抗性提高.本试验结果显示,随着干旱胁迫程度的加剧,叶片总气孔密度、气孔开张比呈先增大后减小的趋势;而气孔开张宽度呈逐渐减小的趋势,可能是气孔密度的增加有助于植物更好地控制呼吸作用,调节体内H2O和CO2的进出,同时脱落酸(ABA)触发保卫细胞释放K+,保卫细胞渗透压和体积的降低导致气孔开张宽度逐渐减小,甚至关闭.与张晓艳等[26]研究发现气孔密度随干早程度的加剧呈先增加后减小结果一致,说明干旱胁迫程度加速了气孔细胞的程序性死亡所造成的[27],与赵殊丽[28]、齐红岩等[29]、高彦萍等[30]的研究气孔密度逐渐增大结果不同,这可能是甘草叶片具有表皮毛、腺体等适应干旱的特征,对干旱逆境胁迫具有交叉适应性,既能提高植株的抗旱能力,又能降低其在干旱环境下所受到的伤害. ...
水分胁迫下不同抗旱类型大豆叶片气孔特性变化研究
1
2007
... 植物抵御干旱胁迫主动适应环境的一个重要调节因素就是气孔调节[21],干旱胁迫会诱导植物的气孔出现微形态的变化,如气孔密度增大[22]、气孔开度减小[23~24]、气孔下陷、气孔器长度和宽度显著减小等[25],以使植物抗性提高.本试验结果显示,随着干旱胁迫程度的加剧,叶片总气孔密度、气孔开张比呈先增大后减小的趋势;而气孔开张宽度呈逐渐减小的趋势,可能是气孔密度的增加有助于植物更好地控制呼吸作用,调节体内H2O和CO2的进出,同时脱落酸(ABA)触发保卫细胞释放K+,保卫细胞渗透压和体积的降低导致气孔开张宽度逐渐减小,甚至关闭.与张晓艳等[26]研究发现气孔密度随干早程度的加剧呈先增加后减小结果一致,说明干旱胁迫程度加速了气孔细胞的程序性死亡所造成的[27],与赵殊丽[28]、齐红岩等[29]、高彦萍等[30]的研究气孔密度逐渐增大结果不同,这可能是甘草叶片具有表皮毛、腺体等适应干旱的特征,对干旱逆境胁迫具有交叉适应性,既能提高植株的抗旱能力,又能降低其在干旱环境下所受到的伤害. ...
鄂尔多斯高原锦鸡儿属植物叶表皮特征及生态适应性
1
2005
... 旱生植物叶表皮及其附属物发生明显变化,如表皮毛、腺毛、瘤状或乳状突起等.杨九艳等[31~32]认为表皮毛反射强光使得叶片降温,免受灼伤,可代替蒸腾作用而间接减少蒸腾用水.宋玉霞等[33]对贺兰山几种旱生灌木叶表皮进行了扫描电镜观察,认为表皮毛的附着可起到防止日灼,对叶表皮细胞和叶肉细胞均起着很大的保护作用,还可以在气孔开放时起保护作用,减少水分的蒸腾.本试验结果显示,在正常供水下,泌腺及其分泌物多呈光滑圆盘状,泌腺在叶片上下表皮分布数量不均,在下表皮分布数量大于上表皮,其中衰老叶泌腺数量显著低于新叶和成熟叶,可能是干旱胁迫导致叶片PCD,继而影响了泌腺发育;随着干旱胁迫程度的加剧叶片上表皮和下表皮腺体密度总数整体上呈增大的趋势,腺体颜色随着干旱胁迫程度的加剧逐渐加深,形状出现不规则褶皱和内陷,3种类型叶片腺毛数量均比对照多,表明甘草叶片表面的腺体特征参与抗旱逆境调节,从而避免甘草植株受到干旱逆境胁迫的伤害.这与Randhawa等[34]研究发现抗性强的品种具有数量更多的腺毛结果一致. ...
鄂尔多斯高原锦鸡儿属植物叶表皮特征及生态适应性
1
2005
... 旱生植物叶表皮及其附属物发生明显变化,如表皮毛、腺毛、瘤状或乳状突起等.杨九艳等[31~32]认为表皮毛反射强光使得叶片降温,免受灼伤,可代替蒸腾作用而间接减少蒸腾用水.宋玉霞等[33]对贺兰山几种旱生灌木叶表皮进行了扫描电镜观察,认为表皮毛的附着可起到防止日灼,对叶表皮细胞和叶肉细胞均起着很大的保护作用,还可以在气孔开放时起保护作用,减少水分的蒸腾.本试验结果显示,在正常供水下,泌腺及其分泌物多呈光滑圆盘状,泌腺在叶片上下表皮分布数量不均,在下表皮分布数量大于上表皮,其中衰老叶泌腺数量显著低于新叶和成熟叶,可能是干旱胁迫导致叶片PCD,继而影响了泌腺发育;随着干旱胁迫程度的加剧叶片上表皮和下表皮腺体密度总数整体上呈增大的趋势,腺体颜色随着干旱胁迫程度的加剧逐渐加深,形状出现不规则褶皱和内陷,3种类型叶片腺毛数量均比对照多,表明甘草叶片表面的腺体特征参与抗旱逆境调节,从而避免甘草植株受到干旱逆境胁迫的伤害.这与Randhawa等[34]研究发现抗性强的品种具有数量更多的腺毛结果一致. ...
5种锦鸡儿属植物渗透调节物质的变化
1
2005
... 旱生植物叶表皮及其附属物发生明显变化,如表皮毛、腺毛、瘤状或乳状突起等.杨九艳等[31~32]认为表皮毛反射强光使得叶片降温,免受灼伤,可代替蒸腾作用而间接减少蒸腾用水.宋玉霞等[33]对贺兰山几种旱生灌木叶表皮进行了扫描电镜观察,认为表皮毛的附着可起到防止日灼,对叶表皮细胞和叶肉细胞均起着很大的保护作用,还可以在气孔开放时起保护作用,减少水分的蒸腾.本试验结果显示,在正常供水下,泌腺及其分泌物多呈光滑圆盘状,泌腺在叶片上下表皮分布数量不均,在下表皮分布数量大于上表皮,其中衰老叶泌腺数量显著低于新叶和成熟叶,可能是干旱胁迫导致叶片PCD,继而影响了泌腺发育;随着干旱胁迫程度的加剧叶片上表皮和下表皮腺体密度总数整体上呈增大的趋势,腺体颜色随着干旱胁迫程度的加剧逐渐加深,形状出现不规则褶皱和内陷,3种类型叶片腺毛数量均比对照多,表明甘草叶片表面的腺体特征参与抗旱逆境调节,从而避免甘草植株受到干旱逆境胁迫的伤害.这与Randhawa等[34]研究发现抗性强的品种具有数量更多的腺毛结果一致. ...
5种锦鸡儿属植物渗透调节物质的变化
1
2005
... 旱生植物叶表皮及其附属物发生明显变化,如表皮毛、腺毛、瘤状或乳状突起等.杨九艳等[31~32]认为表皮毛反射强光使得叶片降温,免受灼伤,可代替蒸腾作用而间接减少蒸腾用水.宋玉霞等[33]对贺兰山几种旱生灌木叶表皮进行了扫描电镜观察,认为表皮毛的附着可起到防止日灼,对叶表皮细胞和叶肉细胞均起着很大的保护作用,还可以在气孔开放时起保护作用,减少水分的蒸腾.本试验结果显示,在正常供水下,泌腺及其分泌物多呈光滑圆盘状,泌腺在叶片上下表皮分布数量不均,在下表皮分布数量大于上表皮,其中衰老叶泌腺数量显著低于新叶和成熟叶,可能是干旱胁迫导致叶片PCD,继而影响了泌腺发育;随着干旱胁迫程度的加剧叶片上表皮和下表皮腺体密度总数整体上呈增大的趋势,腺体颜色随着干旱胁迫程度的加剧逐渐加深,形状出现不规则褶皱和内陷,3种类型叶片腺毛数量均比对照多,表明甘草叶片表面的腺体特征参与抗旱逆境调节,从而避免甘草植株受到干旱逆境胁迫的伤害.这与Randhawa等[34]研究发现抗性强的品种具有数量更多的腺毛结果一致. ...
贺兰山15种旱生灌木叶表皮扫描电镜观察
1
2003
... 旱生植物叶表皮及其附属物发生明显变化,如表皮毛、腺毛、瘤状或乳状突起等.杨九艳等[31~32]认为表皮毛反射强光使得叶片降温,免受灼伤,可代替蒸腾作用而间接减少蒸腾用水.宋玉霞等[33]对贺兰山几种旱生灌木叶表皮进行了扫描电镜观察,认为表皮毛的附着可起到防止日灼,对叶表皮细胞和叶肉细胞均起着很大的保护作用,还可以在气孔开放时起保护作用,减少水分的蒸腾.本试验结果显示,在正常供水下,泌腺及其分泌物多呈光滑圆盘状,泌腺在叶片上下表皮分布数量不均,在下表皮分布数量大于上表皮,其中衰老叶泌腺数量显著低于新叶和成熟叶,可能是干旱胁迫导致叶片PCD,继而影响了泌腺发育;随着干旱胁迫程度的加剧叶片上表皮和下表皮腺体密度总数整体上呈增大的趋势,腺体颜色随着干旱胁迫程度的加剧逐渐加深,形状出现不规则褶皱和内陷,3种类型叶片腺毛数量均比对照多,表明甘草叶片表面的腺体特征参与抗旱逆境调节,从而避免甘草植株受到干旱逆境胁迫的伤害.这与Randhawa等[34]研究发现抗性强的品种具有数量更多的腺毛结果一致. ...
贺兰山15种旱生灌木叶表皮扫描电镜观察
1
2003
... 旱生植物叶表皮及其附属物发生明显变化,如表皮毛、腺毛、瘤状或乳状突起等.杨九艳等[31~32]认为表皮毛反射强光使得叶片降温,免受灼伤,可代替蒸腾作用而间接减少蒸腾用水.宋玉霞等[33]对贺兰山几种旱生灌木叶表皮进行了扫描电镜观察,认为表皮毛的附着可起到防止日灼,对叶表皮细胞和叶肉细胞均起着很大的保护作用,还可以在气孔开放时起保护作用,减少水分的蒸腾.本试验结果显示,在正常供水下,泌腺及其分泌物多呈光滑圆盘状,泌腺在叶片上下表皮分布数量不均,在下表皮分布数量大于上表皮,其中衰老叶泌腺数量显著低于新叶和成熟叶,可能是干旱胁迫导致叶片PCD,继而影响了泌腺发育;随着干旱胁迫程度的加剧叶片上表皮和下表皮腺体密度总数整体上呈增大的趋势,腺体颜色随着干旱胁迫程度的加剧逐渐加深,形状出现不规则褶皱和内陷,3种类型叶片腺毛数量均比对照多,表明甘草叶片表面的腺体特征参与抗旱逆境调节,从而避免甘草植株受到干旱逆境胁迫的伤害.这与Randhawa等[34]研究发现抗性强的品种具有数量更多的腺毛结果一致. ...
Comparative assessment of density of glandular hairs,population and size of aperture of stomata in resistant and susceptible cultivars of chickpea to Ascochyta blight disease
1
2009
... 旱生植物叶表皮及其附属物发生明显变化,如表皮毛、腺毛、瘤状或乳状突起等.杨九艳等[31~32]认为表皮毛反射强光使得叶片降温,免受灼伤,可代替蒸腾作用而间接减少蒸腾用水.宋玉霞等[33]对贺兰山几种旱生灌木叶表皮进行了扫描电镜观察,认为表皮毛的附着可起到防止日灼,对叶表皮细胞和叶肉细胞均起着很大的保护作用,还可以在气孔开放时起保护作用,减少水分的蒸腾.本试验结果显示,在正常供水下,泌腺及其分泌物多呈光滑圆盘状,泌腺在叶片上下表皮分布数量不均,在下表皮分布数量大于上表皮,其中衰老叶泌腺数量显著低于新叶和成熟叶,可能是干旱胁迫导致叶片PCD,继而影响了泌腺发育;随着干旱胁迫程度的加剧叶片上表皮和下表皮腺体密度总数整体上呈增大的趋势,腺体颜色随着干旱胁迫程度的加剧逐渐加深,形状出现不规则褶皱和内陷,3种类型叶片腺毛数量均比对照多,表明甘草叶片表面的腺体特征参与抗旱逆境调节,从而避免甘草植株受到干旱逆境胁迫的伤害.这与Randhawa等[34]研究发现抗性强的品种具有数量更多的腺毛结果一致. ...
Photosynthetic limitations in response to water stress and recovery in Mediterranean plants with different growth forms
1
2007
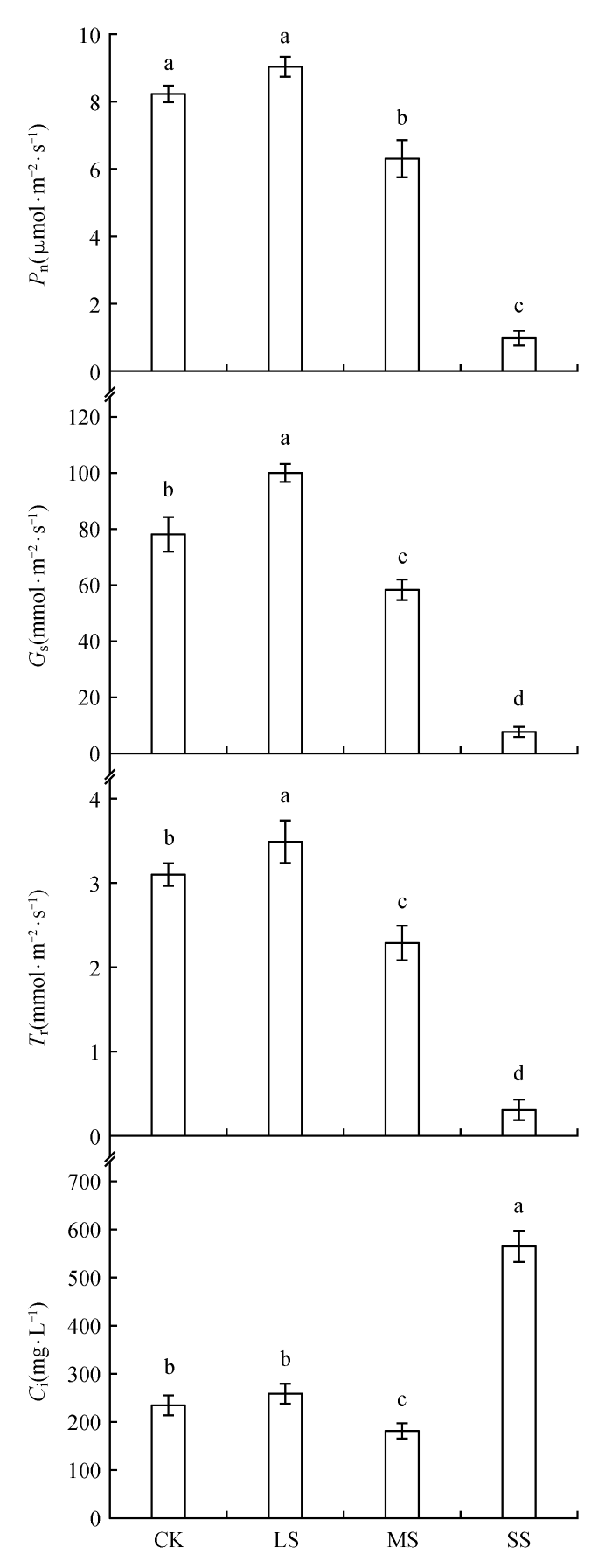
... 干旱胁迫对植物光合作用的影响范围很广,从限制CO2向叶绿体的扩散,限制茎和根产生的激素介导的气孔开放,到限制CO2向叶肉运输,以及改变叶片中光化学反应和碳代谢过程.这些影响随着胁迫的强度和持续时间以及叶片的年龄(老叶片更容易受到干旱的影响,积累更多的盐分)和植物种类而变化[35].近年来的研究表明,在快速或轻度干旱条件下,气孔限制是光合作用下降的主要原因,而缓慢干旱、中度干旱或严重干旱条件下,光合作用主要受非气孔因素限制.植物受旱后,叶片光合作用受抑制的主要原因都有从气孔因素向非气孔因素转变的过程,其转变时间因植物的抗旱能力、干旱程度及施加方式等因素而异.在干旱胁迫下,植物吸水受阻导致体内水分缺乏,蒸腾速率会逐渐减缓,以维持植物基本代谢活动,这是植物抵御干旱胁迫的防御机制[36],气孔导度是蒸腾速率大幅度下降的主要原因.研究表明Pn与Gs和Tr呈显著正相关,是由于气孔在植物光合气体交换中影响着Pn和Gs的过程,其调节能力是植物适应水分逆境的主要措施之一[37].Valladares等[38]研究发现在小麦开花后,小麦叶片净光合速率随干旱胁迫程度的加剧呈先增大再减小的趋势.本试验结果显示随着干旱胁迫程度的加剧,甘草幼苗叶片的Pn、Tr、和Gs都呈现先升高后降低的趋势;在轻度干旱胁迫时,Pn、Tr、和Gs显著高于CK,这可能是甘草叶片表面气孔影响所致.而Ci呈逐渐上升的趋势,在重度胁迫时迅速增高,这与Y.Ohashi[39]等研究结果一致,表明在重度干旱胁迫下甘草幼苗失去抗旱能力,以及限制甘草幼苗光合作用的因素是由非气孔因素导致的. ...
水分胁迫对不同抗旱性砧木嫁接番茄生长发育及水气交换参数的影响
1
2017
... 干旱胁迫对植物光合作用的影响范围很广,从限制CO2向叶绿体的扩散,限制茎和根产生的激素介导的气孔开放,到限制CO2向叶肉运输,以及改变叶片中光化学反应和碳代谢过程.这些影响随着胁迫的强度和持续时间以及叶片的年龄(老叶片更容易受到干旱的影响,积累更多的盐分)和植物种类而变化[35].近年来的研究表明,在快速或轻度干旱条件下,气孔限制是光合作用下降的主要原因,而缓慢干旱、中度干旱或严重干旱条件下,光合作用主要受非气孔因素限制.植物受旱后,叶片光合作用受抑制的主要原因都有从气孔因素向非气孔因素转变的过程,其转变时间因植物的抗旱能力、干旱程度及施加方式等因素而异.在干旱胁迫下,植物吸水受阻导致体内水分缺乏,蒸腾速率会逐渐减缓,以维持植物基本代谢活动,这是植物抵御干旱胁迫的防御机制[36],气孔导度是蒸腾速率大幅度下降的主要原因.研究表明Pn与Gs和Tr呈显著正相关,是由于气孔在植物光合气体交换中影响着Pn和Gs的过程,其调节能力是植物适应水分逆境的主要措施之一[37].Valladares等[38]研究发现在小麦开花后,小麦叶片净光合速率随干旱胁迫程度的加剧呈先增大再减小的趋势.本试验结果显示随着干旱胁迫程度的加剧,甘草幼苗叶片的Pn、Tr、和Gs都呈现先升高后降低的趋势;在轻度干旱胁迫时,Pn、Tr、和Gs显著高于CK,这可能是甘草叶片表面气孔影响所致.而Ci呈逐渐上升的趋势,在重度胁迫时迅速增高,这与Y.Ohashi[39]等研究结果一致,表明在重度干旱胁迫下甘草幼苗失去抗旱能力,以及限制甘草幼苗光合作用的因素是由非气孔因素导致的. ...
水分胁迫对不同抗旱性砧木嫁接番茄生长发育及水气交换参数的影响
1
2017
... 干旱胁迫对植物光合作用的影响范围很广,从限制CO2向叶绿体的扩散,限制茎和根产生的激素介导的气孔开放,到限制CO2向叶肉运输,以及改变叶片中光化学反应和碳代谢过程.这些影响随着胁迫的强度和持续时间以及叶片的年龄(老叶片更容易受到干旱的影响,积累更多的盐分)和植物种类而变化[35].近年来的研究表明,在快速或轻度干旱条件下,气孔限制是光合作用下降的主要原因,而缓慢干旱、中度干旱或严重干旱条件下,光合作用主要受非气孔因素限制.植物受旱后,叶片光合作用受抑制的主要原因都有从气孔因素向非气孔因素转变的过程,其转变时间因植物的抗旱能力、干旱程度及施加方式等因素而异.在干旱胁迫下,植物吸水受阻导致体内水分缺乏,蒸腾速率会逐渐减缓,以维持植物基本代谢活动,这是植物抵御干旱胁迫的防御机制[36],气孔导度是蒸腾速率大幅度下降的主要原因.研究表明Pn与Gs和Tr呈显著正相关,是由于气孔在植物光合气体交换中影响着Pn和Gs的过程,其调节能力是植物适应水分逆境的主要措施之一[37].Valladares等[38]研究发现在小麦开花后,小麦叶片净光合速率随干旱胁迫程度的加剧呈先增大再减小的趋势.本试验结果显示随着干旱胁迫程度的加剧,甘草幼苗叶片的Pn、Tr、和Gs都呈现先升高后降低的趋势;在轻度干旱胁迫时,Pn、Tr、和Gs显著高于CK,这可能是甘草叶片表面气孔影响所致.而Ci呈逐渐上升的趋势,在重度胁迫时迅速增高,这与Y.Ohashi[39]等研究结果一致,表明在重度干旱胁迫下甘草幼苗失去抗旱能力,以及限制甘草幼苗光合作用的因素是由非气孔因素导致的. ...
水分胁迫对柑橘光合特性的影响
1
2010
... 干旱胁迫对植物光合作用的影响范围很广,从限制CO2向叶绿体的扩散,限制茎和根产生的激素介导的气孔开放,到限制CO2向叶肉运输,以及改变叶片中光化学反应和碳代谢过程.这些影响随着胁迫的强度和持续时间以及叶片的年龄(老叶片更容易受到干旱的影响,积累更多的盐分)和植物种类而变化[35].近年来的研究表明,在快速或轻度干旱条件下,气孔限制是光合作用下降的主要原因,而缓慢干旱、中度干旱或严重干旱条件下,光合作用主要受非气孔因素限制.植物受旱后,叶片光合作用受抑制的主要原因都有从气孔因素向非气孔因素转变的过程,其转变时间因植物的抗旱能力、干旱程度及施加方式等因素而异.在干旱胁迫下,植物吸水受阻导致体内水分缺乏,蒸腾速率会逐渐减缓,以维持植物基本代谢活动,这是植物抵御干旱胁迫的防御机制[36],气孔导度是蒸腾速率大幅度下降的主要原因.研究表明Pn与Gs和Tr呈显著正相关,是由于气孔在植物光合气体交换中影响着Pn和Gs的过程,其调节能力是植物适应水分逆境的主要措施之一[37].Valladares等[38]研究发现在小麦开花后,小麦叶片净光合速率随干旱胁迫程度的加剧呈先增大再减小的趋势.本试验结果显示随着干旱胁迫程度的加剧,甘草幼苗叶片的Pn、Tr、和Gs都呈现先升高后降低的趋势;在轻度干旱胁迫时,Pn、Tr、和Gs显著高于CK,这可能是甘草叶片表面气孔影响所致.而Ci呈逐渐上升的趋势,在重度胁迫时迅速增高,这与Y.Ohashi[39]等研究结果一致,表明在重度干旱胁迫下甘草幼苗失去抗旱能力,以及限制甘草幼苗光合作用的因素是由非气孔因素导致的. ...
水分胁迫对柑橘光合特性的影响
1
2010
... 干旱胁迫对植物光合作用的影响范围很广,从限制CO2向叶绿体的扩散,限制茎和根产生的激素介导的气孔开放,到限制CO2向叶肉运输,以及改变叶片中光化学反应和碳代谢过程.这些影响随着胁迫的强度和持续时间以及叶片的年龄(老叶片更容易受到干旱的影响,积累更多的盐分)和植物种类而变化[35].近年来的研究表明,在快速或轻度干旱条件下,气孔限制是光合作用下降的主要原因,而缓慢干旱、中度干旱或严重干旱条件下,光合作用主要受非气孔因素限制.植物受旱后,叶片光合作用受抑制的主要原因都有从气孔因素向非气孔因素转变的过程,其转变时间因植物的抗旱能力、干旱程度及施加方式等因素而异.在干旱胁迫下,植物吸水受阻导致体内水分缺乏,蒸腾速率会逐渐减缓,以维持植物基本代谢活动,这是植物抵御干旱胁迫的防御机制[36],气孔导度是蒸腾速率大幅度下降的主要原因.研究表明Pn与Gs和Tr呈显著正相关,是由于气孔在植物光合气体交换中影响着Pn和Gs的过程,其调节能力是植物适应水分逆境的主要措施之一[37].Valladares等[38]研究发现在小麦开花后,小麦叶片净光合速率随干旱胁迫程度的加剧呈先增大再减小的趋势.本试验结果显示随着干旱胁迫程度的加剧,甘草幼苗叶片的Pn、Tr、和Gs都呈现先升高后降低的趋势;在轻度干旱胁迫时,Pn、Tr、和Gs显著高于CK,这可能是甘草叶片表面气孔影响所致.而Ci呈逐渐上升的趋势,在重度胁迫时迅速增高,这与Y.Ohashi[39]等研究结果一致,表明在重度干旱胁迫下甘草幼苗失去抗旱能力,以及限制甘草幼苗光合作用的因素是由非气孔因素导致的. ...
Drought can be more critical in the shade than in the sun:a field study of carbon gain and photo-inhibition in a Californian shrub during a dry El Ni?o year
1
2002
... 干旱胁迫对植物光合作用的影响范围很广,从限制CO2向叶绿体的扩散,限制茎和根产生的激素介导的气孔开放,到限制CO2向叶肉运输,以及改变叶片中光化学反应和碳代谢过程.这些影响随着胁迫的强度和持续时间以及叶片的年龄(老叶片更容易受到干旱的影响,积累更多的盐分)和植物种类而变化[35].近年来的研究表明,在快速或轻度干旱条件下,气孔限制是光合作用下降的主要原因,而缓慢干旱、中度干旱或严重干旱条件下,光合作用主要受非气孔因素限制.植物受旱后,叶片光合作用受抑制的主要原因都有从气孔因素向非气孔因素转变的过程,其转变时间因植物的抗旱能力、干旱程度及施加方式等因素而异.在干旱胁迫下,植物吸水受阻导致体内水分缺乏,蒸腾速率会逐渐减缓,以维持植物基本代谢活动,这是植物抵御干旱胁迫的防御机制[36],气孔导度是蒸腾速率大幅度下降的主要原因.研究表明Pn与Gs和Tr呈显著正相关,是由于气孔在植物光合气体交换中影响着Pn和Gs的过程,其调节能力是植物适应水分逆境的主要措施之一[37].Valladares等[38]研究发现在小麦开花后,小麦叶片净光合速率随干旱胁迫程度的加剧呈先增大再减小的趋势.本试验结果显示随着干旱胁迫程度的加剧,甘草幼苗叶片的Pn、Tr、和Gs都呈现先升高后降低的趋势;在轻度干旱胁迫时,Pn、Tr、和Gs显著高于CK,这可能是甘草叶片表面气孔影响所致.而Ci呈逐渐上升的趋势,在重度胁迫时迅速增高,这与Y.Ohashi[39]等研究结果一致,表明在重度干旱胁迫下甘草幼苗失去抗旱能力,以及限制甘草幼苗光合作用的因素是由非气孔因素导致的. ...
Effects of drought stress on photosynthetic gas exchange,chlorophyll fluorescence and stem diameter of soybean plants
1
2006
... 干旱胁迫对植物光合作用的影响范围很广,从限制CO2向叶绿体的扩散,限制茎和根产生的激素介导的气孔开放,到限制CO2向叶肉运输,以及改变叶片中光化学反应和碳代谢过程.这些影响随着胁迫的强度和持续时间以及叶片的年龄(老叶片更容易受到干旱的影响,积累更多的盐分)和植物种类而变化[35].近年来的研究表明,在快速或轻度干旱条件下,气孔限制是光合作用下降的主要原因,而缓慢干旱、中度干旱或严重干旱条件下,光合作用主要受非气孔因素限制.植物受旱后,叶片光合作用受抑制的主要原因都有从气孔因素向非气孔因素转变的过程,其转变时间因植物的抗旱能力、干旱程度及施加方式等因素而异.在干旱胁迫下,植物吸水受阻导致体内水分缺乏,蒸腾速率会逐渐减缓,以维持植物基本代谢活动,这是植物抵御干旱胁迫的防御机制[36],气孔导度是蒸腾速率大幅度下降的主要原因.研究表明Pn与Gs和Tr呈显著正相关,是由于气孔在植物光合气体交换中影响着Pn和Gs的过程,其调节能力是植物适应水分逆境的主要措施之一[37].Valladares等[38]研究发现在小麦开花后,小麦叶片净光合速率随干旱胁迫程度的加剧呈先增大再减小的趋势.本试验结果显示随着干旱胁迫程度的加剧,甘草幼苗叶片的Pn、Tr、和Gs都呈现先升高后降低的趋势;在轻度干旱胁迫时,Pn、Tr、和Gs显著高于CK,这可能是甘草叶片表面气孔影响所致.而Ci呈逐渐上升的趋势,在重度胁迫时迅速增高,这与Y.Ohashi[39]等研究结果一致,表明在重度干旱胁迫下甘草幼苗失去抗旱能力,以及限制甘草幼苗光合作用的因素是由非气孔因素导致的. ...





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}